BIOACID II – Konsortium 4: Auswirkungen der Ozeanversauerung in einem sich erwärmenden Klima auf Interaktionen zwischen den Arten an den Verteilungsgrenzen: Mechanismen und Konsequenzen auf Ökosystem-Ebene
Leitung: Dr. Felix Christopher Mark, (AWI, Bremerhaven)
Die Versauerung der Meere ist ein zusätzlicher Stressfaktor, der sich parallel zur laufenden Klimaerwärmung entwickelt. Zukünftige Auswirkungen der Ozeanversauerung auf Organismen und Ökosysteme werden voraussichtlich in den Polargebieten aufgrund erhöhter CO2-Löslichkeit in kaltem Wasser und Körperflüssigkeiten und aufgrund der gleichzeitigen Belastung der Organismen durch die starke Erwärmung am größten sein. Gleichzeitig sind thermische Toleranzfenster enger und damit die Empfindlichkeiten gegenüber kombinierten Stress-Effekten wahrscheinlich in kälte-adaptierten polaren Arten im Vergleich zu gemäßigten Arten höher. Der erwartete Anstieg der Kohlendioxid- Konzentration und der Temperatur in den Ozeanen (800-1.000 μatm bzw. 1-2°C bis zum Jahr 2100 (IPCC, 2007)) erweist sich somit als besonders bedrohlich für arktische Ökosysteme.
Mit der Erwärmung der Ozeane bewegen sich die Fischbestände mit den Wassermassen ihrer bevorzugten Temperaturen, um innerhalb eines physiologisch optimalen Temperaturbereichs zu bleiben, vorausgesetzt, dass weitere Faktoren wie die Verfügbarkeit von Nahrungsmitteln und der Wettbewerb mit anderen Arten dies zulassen. Dies ist bereits für mehrere Fischarten in der Nordsee nachgewiesen worden, die sich mit einer Geschwindigkeit von ungefähr 12 km pro Dekade nach Norden bewegt haben (Drinkwater, 2009; Perry et al., 2005). Als Reaktion auf diesen Erwärmungstrend hat der nordarktische Bestand an Kabeljau (Gadus morhua) ebenfalls die Laichplätze in den Norden (Sundby und Nakken, 2008) verschoben und seine Reichweite bis in die Barentssee (Drinkwater, 2009) erweitert. Für den größten Teil des Jahres werden jugendliche Kabeljaue jetzt häufig in den Küstengewässern vor Spitzbergen (Olsen et al., 2010) angetroffen, was zu einem starken Wettbewerb mit dem einheimischen Polardorsch (Boreogadus saida) führt. Polardorsch ist eine dominante Fischart auf dem arktischen Schelf und gilt als eine Schlüsselspezies in arktischen Ökosystemen. Es ist jedoch nicht klar, wie diese beiden Arten auf lange Sicht interagieren werden und ob die Artenzusammensetzung des Ökosystems auf die Änderungen des Einzugsgebietes des Atlantischen G. morhua in der Arktis reagieren wird (Renaud et al., 2011).
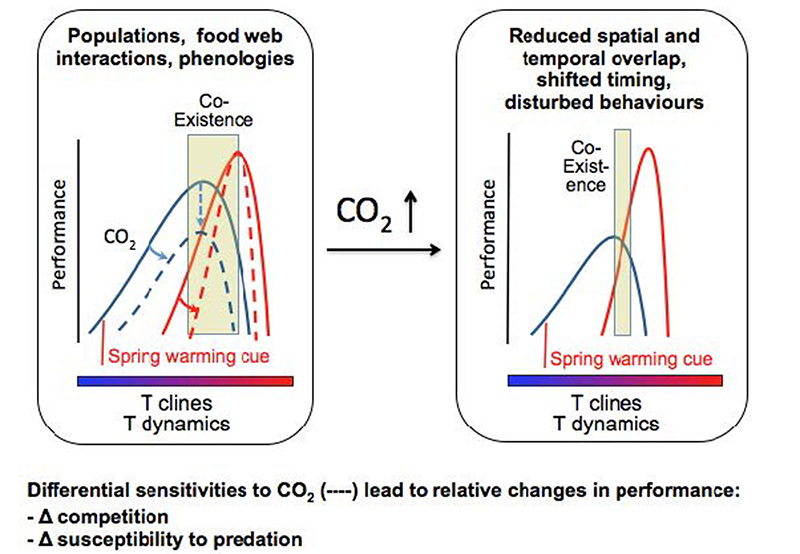
Betrachtung des Ökosystems hinsichtlich der Beeinflussung der Interaktionen zwischen den Arten durch die synergistischen Effekte der veränderten Temperatur, des CO2- und Hypoxie-Niveaus, aufbauend auf einem mechanistischen Verständnis. Die Integration von CO2-Empfindlichkeit auf einer thermischen Matrix, definiert von den Grundsätzen der Sauerstoff- und Kapazitätsaufbau-begrenzten thermischen Toleranz, betont, wie Spezies-spezifische Empfindlichkeiten und ihre minimalen Leistungsniveaus Einfluss nehmen auf das Fenster der zeitlichen und räumlichen Überschneidungen, die den emperaturbereich von Koexistenz sowie der Veränderungen in relativer Leistung definieren (modifiziert nach Pörtner und Farrell, 2008; Pörtner, 2010).
Die Auswirkungen der Ozeanerwärmung auf Fischverbreitung und Physiologie wurden zu einem gewissen Grad für den Kabeljau untersucht und dokumentiert (z.B. Colosimo et al., 2003; Lannig et al., 2003; Loeng und Drinkwater, 2007; Mueter et al., 2009; Pörtner et al., 2008; Pörtner et al., 2001; Rose, 2005) und in geringerem Maße auch für den Polardorsch (Gjosaeter und Ajiad, 1994; Graham und Hop, 1995; Hop und Graham, 1995; Nahrgang et al., 2010). Allerdings ist nur sehr wenig darüber bekannt, wie die Physiologie oder Verteilung dieser Arten durch die zusätzlichen Effekte von Hyperkapnie verändert werden. Außerdem fehlen Daten darüber, wie Hyperkapnie die Interaktionen zwischen den Arten, die bereits von der Erwärmung betroffen sind, ändern wird. Mit Kabeljau und Polardorsch als Modellarten schlagen wir deshalb vor, die Wettbewerbs-Wechselwirkung zwischen zwei derzeit koexistierenden arktischen Fischarten unter der kombinierten Wirkung von Ozeanversauerung und Erwärmung (Ocean Acidification and Warming, OAW) in einem Ökosystem der Arktis, dem Kongsfjord in Spitzbergen, zu studieren. In dieser Fallstudie wollen wir Physiologie, Verhalten und Lebenszyklus der Arten Rechnung tragen und die Hypothese, dass diese Arten unterschiedlich von OAW betroffen sein werden, untersuchen, mit der Folge zukünftiger Veränderungen in ihrer Wettbewerbs-Interaktion. In den Küstengewässern vor Spitzbergen findet Kabeljau sich an der nördlichen Grenze seiner geographischen Verteilung und damit am unteren Ende seines thermischen Toleranzbereichs. Das Gegenteil gilt für Polardorsch. Spitzbergen markiert die südliche Grenze seines Verbreitungsgebietes und das obere Ende seines thermischen Toleranzbereichs. Dies bedeutet, dass innerhalb des Bereichs ihrer Koexistenz um Spitzbergen, Kabeljau und Polardorsch sowohl energetisch begrenzt sind und ihre physiologische Leistung sehr anfällig für weitere Stressoren sein sollte. Die zunehmende Konzentration von Kohlendioxid ist eindeutig einer von ihnen (Pörtner, 2010). Aus dieser Perspektive ist es unsere Ansicht, dass die Versauerung der Ozeane ein entscheidender Faktor für die Steuerung der Interaktion der koexistierender Arten sein kann und dass wahrscheinlich das Ergebnis ihrer unterschiedlichen Fähigkeiten, OAW zu konfrontieren, bestimmt, ob sie z.B. verdrängt oder auf einen weniger günstigen Lebensraum beschränkt werden (Abb. 4.1; Abrams et al., 2008; Matthews et al., 2010).
Für eine ganzheitliche und integrative Analyse der OAW ist es wichtig, die unterschiedlichen Befindlichkeiten und Empfindlichkeiten der verschiedenen Lebensphasen der beiden Arten sowie deren Wechselwirkung mit ihren Beuteorganismen und damit ihre Abhängigkeit von der Nahrungskette zu berücksichtigen. Frühere Untersuchungen über die Auswirkungen des erhöhten CO2-Gehalts auf Meeresfische führten zu der allgemeinen Vorstellung, dass sie aufgrund ihrer mächtigen Mechanismen der Ionen-Regelung für die Versauerung der Ozeane nicht besonders anfällig sind. Erwachsene Kabeljaue sind in der Lage, Säure-Basen-Störungen (Larsen et al., 1997) zu kompensieren und zeigen keine wesentlichen Auswirkungen von Hyperkapnie auf die physiologische Leistung (Melzner et al., 2009a). Allerdings zeigten Studien an verschiedenen Spezies chronische Auswirkungen von Umwelt-Hyperkapnie auf Jungfische (Moran und Stottrup, 2010), und eine Empfindlichkeit der Eier und frühen Lebensstadien, die höher war als bei Erwachsenen. Es muss jedoch erwähnt werden, dass alle diese Beobachtungen für die CO2-Konzentrationen jenseits realistischer Szenarien für die Versauerung der Ozeane erzielt wurden (Ishimatsu, 2005; Kikkawa et al., 2004). Die resultierende Ansicht, dass Fische weitgehend unempfindlich gegenüber Ozeanversauerung sind, mag daher verfrüht sein.
In der Tat zeigten neuere Befunde Verhaltensstörungen bei Fischen in tropischen Korallenriffen, die Hyperkapnie-Niveaus gemäß Ozeanversauerungs-Szenarien ausgesetzt waren (Munday et al, 2009c), und ebenfalls Änderungen in der Larvenentwicklung von Kabeljau (Frommel et al., im Druck) und Hering (Franke und Clemmesen, 2011). Außerdem wurde die Hypothese, dass die Empfindlichkeit gegenüber thermischen Extremen unter den projizierten Ozeanversauerungs-Niveaus erhöht wird, in Korallenfischen bestätigt (Nilsson et al., 2009; Pörtner und Farrell, 2008). Im Allgemeinen kann die Empfindlichkeit der Fische auf die Versauerung der Ozeane bei extremen Temperaturen höher sein, eine Hypothese, die mit Fischen aus verschiedenen Klimazonen getestet werden sollte. Gleichzeitig zeigt die Allometrie der thermischen Einschränkung, dass Toleranz gegenüber extremen Temperaturen über Lebensphasen hinweg variiert und mit zunehmender Körpergröße abnimmt (z.B. Pörtner et al., 2008; Pörtner und Knust, 2007; Storch et al., 2011). Dementsprechend kann die Empfindlichkeit gegenüber OAW auch während der Ontogenese variieren, wie jüngst in Krebstieren beobachtet (Schiffer et al., in Vorb.). Die Konsequenz dieser Beobachtungen für die Larvenrekrutierung bleibt somit weiterhin zu erkunden. Diese fragmentierte Wissensbasis erfordert somit eindeutig ein besseres Verständnis davon, wie die Belastung in kombinierten CO2– und Temperatur-Szenarien zur Gestaltung des Fitness-Fensters beiträgt. Dazu gehört auch die Frage, ob alle Lebensphasen ähnlich reagieren, oder ob einige von ihnen mögliche Engpässe für das Überleben der Population darstellen.
Die meisten Lebensphasen von Kabeljau und Polardorsch ernähren sich von Plankton-Organismen (Levasseur et al., 1994; Lowry und Frost, 1981; Renaud et al., 2011). Ihre Verfügbarkeit und die ernährungsphysiologische Qualität sind ein sehr wichtiger Nebeneffekt der OAW, vermutlich mit Auswirkungen auf die Entwicklung und Leistung der frühen Lebensstadien von beiden Fischarten. Getrieben durch den Klimawandel, unterziehen Planktongemeinschaften sich gut dokumentierten Veränderungen in Zusammensetzung und räumlicher Weite (Beaugrand et al., 2009), die in einem Bottom-Up-Kontrollprozess die biogeographische Verbreitung der Beutefischbestände beeinflussen (Drinkwater, 2009; Drinkwater, 2006). In dieser Hinsicht ist es von erheblicher Bedeutung, dass von Natur aus invasive Arten eher Generalisten als Spezialisten sind und somit erfolgreicher als die einheimischen Arten bei der Anpassung an Veränderungen in Nahrungszusammensetzung und Angebot sein können (Dukes und Mooney, 1999). In den Küstengewässern von Spitzbergen spiegelt sich dies in der Nahrung von Kabeljau und Polardorsch wider (Renaud et al., 2011), mit einer vielfältigeren Zusammensetzung der Nahrungskette für Kabeljau. Dies könnte sich als vorteilhaft erweisen für die Fitness und das Potenzial des Kabeljaus, sich einem wandelnden Klima anzupassen.
Der Erfolg des Beutefangs und damit des Überlebens hängt auch vom engagierten Verhalten des Raubtieres und seiner Beutetiere ab. Studien mit tropischen Korallenrifffischen zeigten, dass Exposition gegenüber erhöhtem CO2 das Vermeidungsverhalten bei Jugendlichen stark gestört hat und in der Folge zu verstärktem Beutefang führte. Diese Verhaltensänderungen sind vermutlich durch Hyperkapnie-Wirkungen auf das zentrale Nervensystem ausgelöst. Durch die Änderung des Artenverhalten hat Hyperkapnie wahrscheinlich das Potenzial, die ausgewogenen Ernährungsnetz-Interaktionen in diesem besonderen Ökosystem durcheinander zu bringen (Ferrari et al., 2011; Munday et al., 2009a; Munday et al., 2010). Dieser Aspekt ist noch nie bei erwachsenen oder jugendlichen gemäßigten und borealen Fischen untersucht worden und muss daher bei den Interaktionen zwischen Kabeljau und Polardorsch und ihrer Beute in Betracht gezogen werden.
Die Gadidae-Familie umfasst einige der kommerziell wichtigsten Fischarten im Nordatlantik, nämlich Kabeljau, Schellfisch, Pollack und Wittling, und Polardorsch ist ebenfalls das Ziel erheblicher industrieller Fischereiaktivitäten in der Arktis (FAO Fischereistatistik). Der Kabeljau ist derzeit Gegenstand einer intensiven Aquakultur in Norwegen. Es ist daher klar, dass jegliche Veränderung in der Bevölkerungsstruktur, die von OAW verursacht wäre, weitreichende Auswirkungen nicht nur auf das Ökosystem selbst, sondern auch auf die Fischerei hätte. Es wurde ebenfalls vorgeschlagen, dass OAW schädliche Auswirkungen auf die Aquakultur (Guinotte und Fabry, 2008) haben könnte. Die sozio-ökonomischen Folgen eines solchen Szenarios müssen noch erforscht werden.
Allgemeine Zielsetzung
Dieses Konsortium wird untersuchen, wie die kombinierten Auswirkungen von OAW die verschiedenen Lebensphasen der wechselwirkenden Fischarten Gadus morhua und Boreogadus saida und ihrer Beute beeinflussen. Die Stärke des Konsortiums liegt in dem integrativen Ansatz über Ebenen der biologischen Organisation hinweg, vom Genom bis zum Ökosystem. Eng verflochtene Arbeitspakete, die sich auf mehrere gemeinsame Akklimatisierungsexperimente berufen, ermöglichen eine gründliche Analyse der sich vermeintlich verschiebenden Interaktionen zwischen den Arten und ihre Auswirkungen auf Ökosystem- und sozioökonomischer Ebene.
Die Zielsetzung beinhaltet auch die Frage, ob sich OAW unterschiedlich auf die wechselwirkenden Spezies auswirkt aufgrund von verschiedenen physiologischen Optima und Bereiche, ausgedrückt im thermischem Toleranzfenster und den damit verbundenen Leistungskapazitäten und Phenologien der spezifischen Lebensphasen. Entscheidende Mechanismen sowie Ursachen und Wirkungen werden durch die Entschlüsselung der Verbindungen zwischen den biologischen Organisationsebenen, von genomischen, molekularen, zellulären, Einzel- und Populationsebenen zugänglich gemacht. Der Umfang von Akklimatisierung (Physiologie und Verhalten) und Anpassung (Evolution), die gemeinsam die Artenwiderstandsfähigkeit definieren, wird in verschiedenen Lebensphasen untersucht (Eier, Larven, Jugendliche, Erwachsene) und das empfindlichste Element (oder Elemente) identifiziert werden. Funktionelle Determinanten der individuellen Fitness, wie Ionen- und Säure-Basen-Regulation, mitochondrialer Energiestoffwechsel und Immunantwort, werden ebenfalls geprüft werden. Darüber hinaus wird untersucht, wie diese Prozesse sowie die Fitness von Nahrungsqualität und -Verfügbarkeit beeinflusst werden.
Das Konsortium wird sich mit folgenden Fragen beschäftigen:
- Wie wird OAW die Verbreitung von Arten, Wettbewerbsfähigkeit und Verhalten beeinflussen (WP 4.1 und 4.6)?
- Werden Nahrungsnetz-bedingte OAW-Effekte die Anfälligkeit der Arten beeinflussen (WP 4.5, 4.8 und 4.9)?
- Welche Lebensphasen sind am anfälligsten für OAW und tragen am meisten zu Auswirkungen auf Ökosystemebene bei (WP 4.1, 4.4, 4.5, 4.6 und 4.7)? Wie zeigen sich OAW-Auswirkungen auf verschiedenen Ebenen der biologischen Organisation, wie bedingen sie einander und wie sind sie zwischen den einzelnen Ebenen miteinander verknüpft (WP 4.1, 4.2, 4.3, 4.4, 4.5 und 4.7)?
Forschungsansätze
Jedes der neun Arbeitspakete in diesem Konsortium beschäftigt sich mit einer bestimmten Ebene der biologischen Organisation, um ihre Ergebnisse in eine Vorhersage der Reaktionen auf Ökosystem-Ebene und deren sozioökonomische Folgen im Konsortium 5 (AP 5.6) zu integrieren.
Die Experimente werden in verschiedenen Laboren durchgeführt. Am Alfred-Wegener-Institut in Bremerhaven werden erwachsene Fische und Jungfische sowie Copepoden in speziell entworfenen OAW-Akklimatisierungssystemen aufbewahrt, die während BIOACID I verbessert (und zum Teil bereits erworben) wurden. Kulturen von Larven und Jungfischen sowie Pteropoden werden unter Laborbedingungen an der Universität Bergen, Norwegen vorbereitet sowie Kulturen von Jungfischen, Pteropoden und Copepoden am AWIPEV Labor in Ny-Ålesund, Spitzbergen. Fische und Copepoden werden von wild lebenden Populationen mit FS HEINCKE um Spitzbergen und Norwegen in 2012 und 2013 in enger Zusammenarbeit mit unseren norwegischen Projektpartnern in Bergen und Tromsø gefangen. Weiterhin bearbeiten wir Fische und Eier aus der norwegischen Aquakultur und benutzen ebenfalls verschiedene Lebensphasen, die am Sven Lovén Zentrum für Marine Wissenschaften in Kristineberg, Schweden (erwachsener Kabeljau, Eier, Larven) und am Nationalen Kabeljau-Zuchtzentrum in Tromsø (erwachsener Polardorsch, Eier, Larven) kultiviert wurden.
Die Arbeitspakete 4.1, 4.2, 4.3, 4.6 und 4.7 benutzen Jungtiere aus gemeinsamen Inkubationen von juvenilem Kabeljau (Frühjahr 2013) und Polardorsch (Frühjahr 2014) bei verschiedenen Temperatur- pCO2-Kombinationen am Alfred-Wegener-Institut. Diese Inkubationen (organisiert von AP 4.1) werden daher als zentrale Drehscheibe für mehrere Arbeitspakete dienen, mit uneingeschränkten Verbindungen zwischen den Arbeitspaketen und gleichzeitiger Reduzierung des Aufwandes und der Anzahl der pro Inkubation benötigten Individuen.
Um die individuelle Akklimatisierungshistorie für jeden Jungfisch nachzuverfolgen, werden einzelne Exemplare aus jeder Abstammung gekennzeichnet und die Bevölkerungen auf ihre Bandbreite bezüglich Wachstum und sonstiger umweltanpassungs-relevanter physiologischer Leistungen („Phänotyp Gesamtorganismus“, AP 4.1) untersucht, als auch auf ihre genetische Variabilität („Genotypen“, AP 4.3). Die Studien werden auch die Vielfalt der Reaktionen festlegen, aus denen die Auswahl und weitere Anpassungen möglich sind, womit die Auflösung und Genauigkeit der Vorhersage auf Bevölkerungsebene erhöht wird. Die Analyse von Transkriptom-Reaktionen (AP 4.3), zusammen mit Anpassungen des Proteoms (AP 4.2), wird Informationen über Anpassungsmechanismen, die sich auf zellulärer und systemischer Ebene manifestieren, zur Verfügung stellen.
Die Berücksichtigung der Vielfalt innerhalb der Bevölkerung wird die Auflösung und Genauigkeit für die Vorhersage auf der Bevölkerungsebene erhöhen. Solche Vorhersagen erfordern auch die Identifizierung der empfindlichsten Lebensphasen und Übergangsphasen einschließlich Reproduktion, Befruchtungserfolg, Ei- und Larvenentwicklung und Metamorphose von Larven zu Jungtieren (AP 4.4 und 4.5). Artenspezifische Unterschiede und Verschiebungen in den Leistungskennzeichen gestalten die Interaktionen in Nahrungsketten (AP 4.5 und 4.6), so eine Hypothese. CO2 kann solche Prozesse aufgrund von Änderungen der relativen Leistung und des Verhaltens beeinflussen. AP 4.6 wird sich damit beschäftigen, die Verhaltensänderungen bei Jungfischen zu identifizieren und die neuronaler Untermauerung der OAW-Effekte auf das Verhalten mittels MRI- und NMR-Spektroskopie zu erläutern.
Für eine Aufnahme der Nahrungsketten-Effekte integrieren wir auch Copepoden und Pteropoden, die unter den gleichen OAW-Bedingungen wie in AP 4.8 und AP 4.9 aufgezogen wurden, und wir quantifizieren die Beute-Aufnahme, die Fangraten von Jungkabeljau (AP 4.6) sowie mögliche Veränderungen in der Nahrungsqualität (AP 4.8 und 4.9). Zu diesem Zweck werden die Arbeitspakete 4.8, 4.9, 4.4, 4.5 und 4.7 gemeinsame Experimente zu Copepoden und Pteropoden-Leistungen durchführen, um die Reaktionen von Eiern und Fischlarven zu CO2 und Änderungen in der Verfügbarkeit von Nahrung zu ermitteln. Die Experimente werden im Sommer 2013 durchgeführt, bei AWIPEV in Ny-Alesund, im Frühjahr/Sommer 2014, Bergen, Norwegen, und an den AWI Einrichtungen. Die Experimente werden in enger Zusammenarbeit mit unseren externen Partnern durchgeführt. Zur Prüfung der Rolle solcher Phänomene unter Feldbedingungen werden diese Arbeitspakete auch mit dem BIOACID-Konsortium 1 während der gemeinsamen Mesokosmen-Studien (KOSMOS) im Gullmar Fjord, Kristineberg, Schweden, im Frühjahr und Sommer 2013 zusammenarbeiten.
In Zusammenarbeit mit dem Brückenarbeitspaket AP 5.6 in Konsortium 5 werden wir letztendlich die sozio-ökonomischen Auswirkungen auf Fischerei und Aquakultur und ihre gesellschaftliche Relevanz etablieren. Dazu werden wir uns einem internationalen Netzwerk von Wissenschaftlern zur Ozeanversauerung, die bereits die sozioökonomischen Aspekte der OAW untersuchen, verbinden. Das stellt ein weiteres Mittel dar, die Ergebnisse dieses Konsortiums zu extrapolieren und an die Öffentlichkeit weiterzugeben.
Zurück zum wissenschaftlichen Programm BIOACID II